
The Bible plainly teaches that the Flood of Noah's day covered the entire globe. Old Testament writers believed it. Jesus Christ and the Apostle Peter based key doctrines on it as a fact of history.
But this raises many serious questions, such as: How could Noah and his family have cared for 16,000 animals? What did the camivorous animals eat during and after the Flood? Lack of answers to these questions had encouraged theologians in the early 1800's to deny Biblical inerrancy and open the door for full-scale denial in the days of Darwin. Today skeptics make great fun of the whole account, and believers often retreat to local-flood compromises.
This book presents good answers to all these questions, Biologist John Woodmorappe has shown that there are always satisfying answers, even to the hard questions, if we are willing to study and be open-minded. No longer can there be an excuse for theological disbelief or the jokes of scoffers. Noah's Ark: A Feasibility Study ably shores up the foundation of this crucial Scriptural doctrine.
From Chapter 14
Boarding the Ark: The Fallacy of Climatic Barriers
Bats and Reptiles on the Ark
I now focus on the most temperature-sensitive animals on the Ark itself. How could temperature needs of all the animals on the Ark have been met simultaneously? We need to keep this apparent difficulty in perspective by realizing that most animals on the Ark could have tolerated a wide range of temperature. For instance, the comfort zone of cattle and sheep is 5 – 2 1 C (E 1991, p. 115 – 6). Most other captive mammals are comfortable at a housing temperature within range of 15 – 25 C (CCAC 1984, vol. 1, p. 80). Considering modern bats, there is a large overlap in their respective temperature requirements. For instance, tropical bats can be kept at a temperature of 25 – 28 C, with temperate bats down to 22 C (Barnard 1991, p. 9). The key to meeting their thermal needs is to provide a temperature gradient (see below). 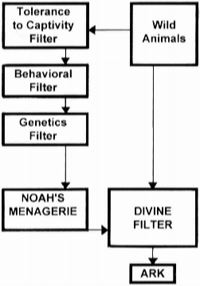 Let us now consider reptiles. If their temperature and photoperiod requirements are not met, their growth will be limited and their feeding will be reduced (Barnard 1991, p. 20). Sometimes they decline and die. However, this decline is over a relatively long period of time (Frye 1991, Vol. 1, p. 161). I had earlier pointed out that most captive animals show an asymptotic decline in death rate with time after the animals have been in captivity. Likewise, the experiments of Licht et al. (1969, p. 481), involving temperature as a variable, show that captive lizards not only die off with an asymptotically-declining death rate with time, but that the survivors appear to tolerate changes in temperatures quite well. If this can be generalized to other reptiles, then it has a practical application in terms of decreasing the reptilian sensitivity to possibly unfavorable temperatures on the Ark. Had reptiles been in captivity prior to their embarkation on the Ark, those individuals that were more sensitive to temperature changes would have already died off before the time came for God to commandeer the reptilian pairs into the Ark. (This is another example of the tolerance-to-captivity filter: Figure. 6). Let us now consider reptiles. If their temperature and photoperiod requirements are not met, their growth will be limited and their feeding will be reduced (Barnard 1991, p. 20). Sometimes they decline and die. However, this decline is over a relatively long period of time (Frye 1991, Vol. 1, p. 161). I had earlier pointed out that most captive animals show an asymptotic decline in death rate with time after the animals have been in captivity. Likewise, the experiments of Licht et al. (1969, p. 481), involving temperature as a variable, show that captive lizards not only die off with an asymptotically-declining death rate with time, but that the survivors appear to tolerate changes in temperatures quite well. If this can be generalized to other reptiles, then it has a practical application in terms of decreasing the reptilian sensitivity to possibly unfavorable temperatures on the Ark. Had reptiles been in captivity prior to their embarkation on the Ark, those individuals that were more sensitive to temperature changes would have already died off before the time came for God to commandeer the reptilian pairs into the Ark. (This is another example of the tolerance-to-captivity filter: Figure. 6).
The exact temperature requirements have not been established for many types of reptiles (Mattison 1987, p. 44). The problem of not knowing the specific thermal preferendum of a reptile is solved by providing a temperature gradient in its cage (Gans and van den Sande 1976, p. 30). This creates a thermally-diversified environment, and gives the reptile an opportunity to choose its temperatures. At the same time, we should not overstate the limitations posed by reptilian thermal requirements, as reptiles are not nearly as fragile as Moore (1983, p. 31) makes them out to be. In reality, most reptiles are robust and easily cared for (Mason et al. 1991, p. 84). Numerous snakes have been successfully maintained without specialized temperature conditions, and at near room temperatures (or slightly warmer), for long periods of time (Murphy and Campbell 1987, p. 167). Furthermore, there is some evidence that reptiles can become acclimated to different climatic conditions (Schwaner 1989, p. 398). Variable Thermal Environment on the Ark.
There are a number of ways that a temperature gradient could have been provided on the Ark for temperature-sensitive animals. For instance, concave mirrors could have been used to concentrate sunlight into certain areas of the Ark. This is feasible because the use of mirrors definitely goes back to early antiquity. For instance, the ancient Chinese knew how to make concave mirrors (CAS 1983, pp. 171 – 2). These have also been used experimentally to study the effects of sunlight when reflected upon reptiles (White 1973, p. 507). To effect a temperature gradient, for example, elongated reptile-bearing cages could have had one bathing in heat and the other end in cold. Post-Flood Origins of Thermal Preferenda.
Up to now I have, for the sake of discussion, accepted the present-day thermal preferenda of reptiles and bats as if they had been valid at the of the Flood. In actuality, these ecological specializations (temperature, photoperiod, etc.) are mostly post-Flood microevolutionary occurrences. One evidence for this is the fact that optimal temperature requirements of iguanid lizards vary according to species (Huey and Kingsolver 1993, 526), and the same holds true for many other reptiles (see Table 1 of Heatwole 1976). Furthermore, for reptiles, thermal preferenda vary within species, according to environment, by season of the year, and according to body mass of the individual reptile (Smith and Ballinger 1994, p. 2066).
|
